Die Koboldmakis
Die Koboldmakis Südostasiens (Gattung Tarsius) gehören zu den kleinsten und ungewöhnlichsten aller heute lebenden Primaten.
In vielerlei Hinsicht sind sie anatomisch eine Mittelform zwischen den Feuchtnasenaffen (Strepsirrhini) und den Eigentlichen Affen (Anthropoidea).
Ihre Ähnlichkeiten mit den Feuchtnasenaffen sind allesamt primitiver Natur: eine nicht verschmolzene Unterkiefersymphyse, Backenzähne mit hohen Höckern, Putzkrallen an ihren zweiten (und dritten) Zehen, mehrere Brustwarzen, und eine zweihörnige Gebärmutter (Uterus bicornis). Ihre Ähnlichkeiten mit höheren Primaten scheinen dagegen abgeleitete Spezialisierungen zu sein, ein Indiz für eine phyletische Beziehung beider Gruppen. Darüber hinaus haben Koboldmakis (Tarsius) viele verschiedene Merkmale, die man bei allen anderen Primaten vergeblich sucht.
Das auffälligste Merkmal der Koboldmakis (Tarsius) sind ihre großen Augen, von denen jedes einzelne tatsächlich größer als das Gehirn ist. Obwohl nachtaktiv, unterscheiden sich Koboldmakis von anderen nachtaktiven Primaten und gleichen mit einer retinalen Fovea eher den tagaktiven Primaten - und das reflektierende Tapetum lucidum - wie man es bei allen Lemuren und Loris, sowie vielen anderen Säugetiergruppen findet - fehlt bei ihnen. Ihre großen Augen werden von einer knöchernen Tasche geschützt, ähnlich wie bei höheren Primaten. Die Nase der Koboldmakis ähnelt ebenfalls höheren Primaten, sowohl außen durch eine nicht befestigte Oberlippe mit einer medianen Falte, als auch intern mit stark reduzierten Nasenmuscheln und durch das Fehlen einer Aussparung des Siebbeins (Os ethmoidale). Wie bei höheren Primaten erfolgt die Blutversorgung des Gehirns durch die innere Halsschlagader (Arteria carotis interna). Der Trommelfellring liegt außerhalb der Gehörbulla und erweitert sich seitlich zu der knöchernen Röhre des äußeren Gehörgangs.

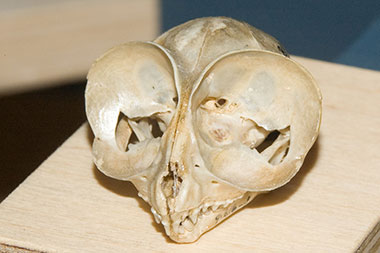

Die Zahnformel der Koboldmakis ist einzigartig unter den Primaten und lautet $\tfrac {2.1.3.3} {1.1.3.3}$, ihre Zähne ähneln jedoch in ihren Gesamtproportionen mit großen oberen mittleren Schneidezähnen und großen Eckzähnen denen der eigentlichen Affen (Anthropoidea). Ihre einfachen Backenzähne mit hohen Höckern auf den oberen Molaren sehen oberflächlich sehr primitiv aus, aber es gibt Indizien dafür, dass sie sich aus einem vormals komplexen Backenzahntyp in ihrer Abstammungslinie entwickelt haben.
Das postcraniale Skelett der Koboldmakis ist in vielen Proportionen auffällig. Die Hände und Füße dieser winzigen Primaten sind relativ groß, was sowohl ihre Fähigkeit, sich perfekt an Stämme und Äste festzuklammern, sowie ihre räuberische Lebensweise reflektiert. Sie haben extrem lange Beine und viele weitere spezifische Anpassungen, die für kraftvolles Springen notwendig sind, einschließlich verschmolzene Schien- und Wadenbeine und eine sehr lange Sprunggelenksregion.
In ihrer Fortpflanzungsphysiologie zeigen Koboldmakis mehrere Ähnlichkeiten mit höheren Primaten. Genau wie Affen, Menschenaffen und Menschen haben sie eine hemochoriale Plazenta (Placenta discoidalis) statt einer epitheliochorialen Plazenta (Placenta diffusa), wie man sie bei Lemuren findet. Außerdem sind ihre Neugeborenen bereits bei der Geburt ziemlich groß. Weibliche Koboldmakis zeigen monatliche Sexualzyklen mit Schwellungen, die an einige Altweltaffen erinnern.

Es gibt 10 Arten von Koboldmakis, die man in die drei Gruppen Sunda-, Philippinen- und Sulawesi-Gruppe aufteilt:
- zur Sunda-Gruppe zählt man den Sunda-Koboldmaki (Tarsius bancanus) auf Borneo, Java und Sumatra,
- zur Philippinen-Gruppe zählt man den Philippinen-Koboldmaki (Tarsius syrichta) auf den Philippinen,
- zur Sulawesi-Gruppe gehören der Sulawesi-Koboldmaki (Tarsius tarsier) auf Sulawesi, der Diana-Koboldmaki (Tarsius dentatus) im Zentrum Sulawesis, der kleine Zwergkoboldmaki (Tarsius pumilus) aus montanen Wäldern, ebenfalls auf Sulawesi, sowie die neu beschriebenen Arten Lariang-Koboldmaki (Tarsius lariang, westlicher Teil Sulawesis), Peleng-Koboldmaki (Tarsius pelengensis, Insel Peleng vor der Ostküste Sulawesis), Sangihe-Koboldmaki (Tarsius sangirensis, von der Insel Sangihe Besar zwischen Sulawesi und Mindanao), Siau-Koboldmaki (Tarsius tumpara, von der Insel Siau zwischen Sulawesi und Mindanao) und Wallace-Koboldmaki (Tarsius wallacei, aus Zentral-Sulawesi).
Der Sunda-Koboldmaki (Tarsius bancanus) und der Sulawesi-Koboldmaki (Tarsius tarsier) scheinen am häufigsten in den unteren Schichten aller Arten von Wäldern vorzukommen, sind aber auch besonders zahlreich in sekundären Wäldern und Buschland anzutreffen. Koboldmakis sind ausschließlich nachtaktiv und verbringen den Tag schlafend im Gras oder auf Kletterpflanzen. Sowohl Sunda-Koboldmaki (Tarsius bancanus) als auch Sulawesi-Koboldmaki (Tarsius tarsier) gehen sehr nahe am Boden auf die Jagd. Sie bewegen sich vor allem durch schnelle Sprünge von bis zu 3 m fort. Auf Sulawesi finden andere Aktivitäten wie etwa Ruhepausen höher in den Baumkronen statt. Der Zwergkoboldmaki (Tarsius pumilus) ist mit seinen gekielten Nägeln zum Festhalten an bemoosten Bäumen recht ungewöhnlich. Koboldmakis sind ausschließlich Fleischfresser, sie machen Jagd auf Insekten, Spinnentiere und kleine Wirbeltiere wie Schlangen und Eidechsen.
Koboldmakis scheinen entweder in Kleinfamilien, bestehend aus einem Paar und deren Nachkommen, oder in einem sozialen System zu leben, in dem das Territorium eines Männchens die kleineren Territorien mehrerer Weibchen überlappt. Aus Studien von in Gefangenschaft lebenden Koboldmakis gibt es Hinweise, dass es zum Teil erhebliche Unterschiede im Sozialverhalten der einzelnen Arten gibt. Sulawesi-Koboldmakis (Tarsius tarsier) leben in Familien, schlafen zusammen, geben komplexe, territoriale Duette am frühen Abend und gehen dann die ganze Nacht gemeinsam auf Nahrungssuche. Sie verjagen andere Koboldmakis tatsächlich aus ihren Territorien. Auf der anderen Seite haben Männchen und Weibchen der Sunda-Koboldmakis (Tarsius bancanus) getrennte, überlappende Territorien und gehen alleine auf Nahrungssuche (Crompton und Andau, 1987). Die Weibchen aller Koboldmakiarten bringen nach einer sechsmonatigen Tragzeit ein einzelnes Junges zur Welt. Die Neugeborenen haben bei der Geburt bereits 30% des Gewichts der Mutter und werden für zwei Monate gesäugt. Für Aufzucht und Pflege des Nachwuchses sind weitgehend die Weibchen verantwortlich. Im Gegensatz zu vielen anderen, monogamen Primatenarten gibt es bei Koboldmakis keine Anzeichen, dass sich die Männchen an der Säuglingspflege beteiligen.
Das Verwandschaftsverhältnis von Koboldmakis zu anderen Primaten, sowohl lebenden als auch fossilen, ist seit vielen Jahren ein Thema und bis heute Gegenstand lebhafter Kontroversen. Auf der Grundlage ihrer vielen primitiven Merkmale, sowie der Morphologie der Chromosomen sind Koboldmakis früher zu den feuchtnasigen Halbaffen gezählt worden. Doch viele Autoren wiesen darauf hin, dass Koboldmakis und höhere Primaten eine genügend große Anzahl von markanten Schädel-, Zahn-, und Fortpflanzungsmerkmalen teilen, die es zu rechtfertigen scheinen, sie in eine eigene Teilordnung (Tarsiiformes) innerhalb der Trockennasenaffen (Haplorhini) einzugruppieren.
Systematik
Unabhängig von der Klassifizierung, der man folgt, ist die evolutionäre Geschichte der Koboldmakis weit davon entfernt, gelöst zu sein, und immer mehr Anzeichen von Unterschieden in der Morphologie und dem Verhalten zwischen den Arten der Koboldmakis verkomplizieren das Bild weiter. In der Vergangenheit haben die meisten Forscher Koboldmakis als eine Gruppe von Halbaffen angesehen, aus denen sich die Eigentlichen Affen (Anthropoidea) entwickelten. Doch die vielen Ähnlichkeiten mit Anthropoiden, wie das Fehlen eines Tapetum lucidum, ihre teilweise postorbitale Schließung und ihre im Verhältnis zur Körpergröße sehr großen Zähne, deuten eher darauf hin, dass sie eine Gruppe von Anthropoiden sind, die anpassungsmäßig wieder zurück in die "Halbaffenzone" gegangen sind (Cartmill, 1980). Vielleicht sind Koboldmakis tatsächlich zwergwüchsige Anthropoiden. Die phyletische Geschichte der Koboldmakis und ihre Ähnlichkeit mit höheren Primaten sind wichtige Themen in der Primatenevolution, die gegenwärtig noch ungelöst sind.
Literatur
Cartmill M. 1980. Morphology, function, and evolution of the anthropoid postorbital septum. In: R. L. Ciochon and A. B. Chiarell, Evolutionary Biology of the New World Monkeys and Continental Drift (Biological Regulation and Development), pp. 243-247. New York: Plenum Press.Aiello L. C. 1986. The relationships of the tarsiformes: A review of the case for the Haplorhini. In: B. Wood, L. Martin, and P. Andrews (eds.) Major Topics in Primate and Human Evolution, pp. 47 - 65.
Crompton R. H, Andau P. M. 1987. Ranging, activity rythms, and sociality in free-ranging Tarsius bancanus: A preliminary report. Int. J. Primatol. 8:43-72. DOI: 10.1007/BF02737113
Gunnell G., Rose K. 2002. Tarsiiformes: Evolutionary History and Adaptation. In: W. C. Hartwig (ed.) The Primate Fossil Record (Cambridge Studies in Biological and Evolutionary Anthropology). Cambridge University Press.
Simons E. L. 2003. The Fossil Record of Tarsier Evolution. In: P. C. Wright, E. Simons, S. Gursky (eds.), Tarsiers: Past, Present, and Future (Rutgers Series in Human Evolution). New Brunswick: Rutgers University Press.
Shekelle M., Groves C., Merker S., Supriatna J. 2008. Tarsius tumpara: A New Tarsier Species from Siau Island, North Sulawesi. Primate Conservation 23: 55-64.
 Volltext-PDF
Volltext-PDFMerker S., Driller C., Dahruddin H., Wirdateti, Sinaga W., Perwitasari-Farajallah D. and Shekelle M. 2010. Tarsius wallacei: A New Tarsier Species from Central Sulawesi Occupies a Discontinuous Range. International Journal of Primatology 31(6):1107-1122.
DOI: 10.1007/s10764-010-9452-0